

How To Draw A Genetic Map
How would you experience if y'all had to be the one to claiming Gregor Mendel's image-shifting laws of inheritance? Yet Thomas Hunt Morgan did exactly this and in the process made gene mapping possible.
In 1911, while studying the chromosome theory of heredity, biologist Thomas Hunt Morgan had a major breakthrough. Morgan occasionally noticed that "linked" traits would divide. Meanwhile, other traits on the same chromosome showed piddling detectable linkage. Morgan considered the prove and proposed that a process of crossing over, or recombination, might explain his results. Specifically, he proposed that the two paired chromosomes could "cross over" to substitution information. Today, we know that recombination does indeed occur during prophase of meiosis (Figure 1), and it creates different combinations of alleles in the gametes that result (i.e., the F1 generation; Effigy two).
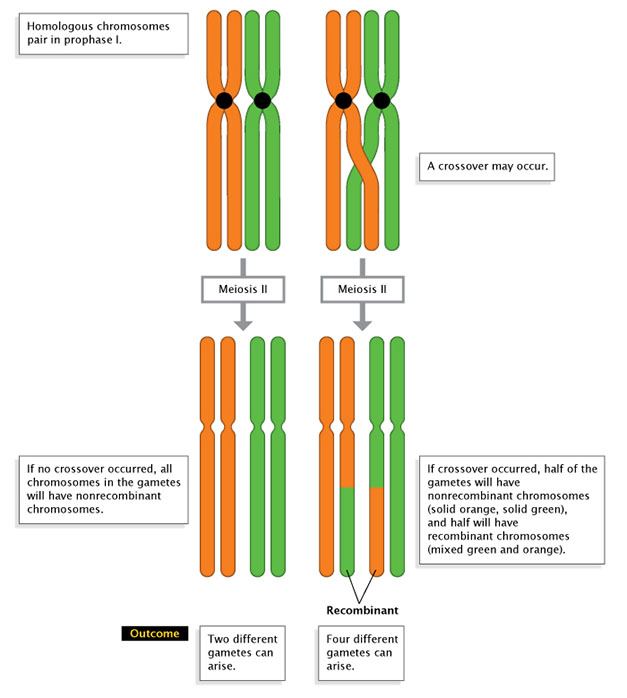
Figure 1: Recombination and gamete product.
A comparing of nonrecombination (left) with recombination (right), shows how recombination affects the style chromosomes are passed into gametes in Meiosis II. On the correct, a single crossover event produces half nonrecombinant gametes and half recombinant gametes.
© 2022 Nature Didactics Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved. ![]()
When proposing the thought of crossing over, Morgan also hypothesized that the frequency of recombination was related to the altitude betwixt the genes on a chromosome, and that the interchange of genetic information broke the linkage between genes. Morgan imagined that genes on chromosomes were similar to pearls on a string (Weiner, 1999); in other words, they were physical objects. The closer two genes were to one another on a chromosome, the greater their chance of existence inherited together. In dissimilarity, genes located farther abroad from one another on the same chromosome were more likely to exist separated during recombination. Therefore, Morgan correctly proposed that the strength of linkage between two genes depends upon the distance between the genes on the chromosome. This proffer became the basis for construction of the earliest maps of the human genome.
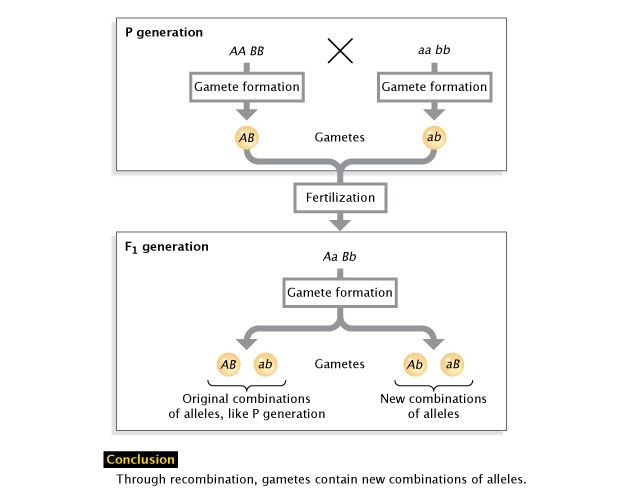
Figure 2: Allele recombination.
Recombination is the sorting of alleles into new combinations. Following the formation of gametes over ii generations shows how recombination tin produce new allelic combinations (lower correct) or stay the same (lower left).
© 2022 Nature Didactics Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved. ![]()
Sturtevant Uses Crossing-Over Information to Construct the Kickoff Genetic Map
Soon later on Morgan presented his hypothesis, Alfred Henry Sturtevant, a 19-year-one-time Columbia University undergraduate who was working with Morgan, realized that if the frequency of crossing over was related to distance, 1 could use this information to map out the genes on a chromosome. Later all, the farther apart two genes were on a chromosome, the more likely information technology was that these genes would separate during recombination. Therefore, as Sturtevant explained it, the "proportion of crossovers could exist used as an index of the distance between any two factors" (Sturtevant, 1913). Collecting a stack of laboratory data, Sturtevant went home and spent almost of the night cartoon the showtime chromosomal linkage map for the genes located on the X chromosome of fruit flies (Weiner, 1999).
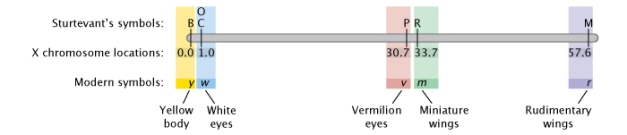
Figure 3: Sturtevant's Drosophila gene map.
In Sturtevant's gene map, six traits are arranged forth a linear chromosome co-ordinate to the relative distance of each from trait B. Traits include xanthous body (B), white eyes (C, O), Vermillion eyes (P), miniature wings (R), and rudimentary wings (M).
© 2022 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, second ed. All rights reserved. ![]()
When creating his map, Sturtevant started by placing six X-linked genes in order. B was a cistron for black torso color. C was a factor that allowed color to appear in the eyes. Flies with the P cistron had vermilion optics instead of the ordinary blood-red, and flies with 2 copies of the recessive O gene had eyes that appeared a shade known as eosin. The R and M factors both affected the wings. Sturtevant placed C and O at the same bespeak because they were completely linked and were always inherited together — in other words, he never saw any bear witness for recombination between C and O. Sturtevant then placed the residuum of the genes in the order shown in Figure 3 (Sturtevant, 1913). Crossover events were tracked past examining the F2 progeny in crosses for "new" phenotypes.

Figure iv: Phenotypes used in Sturtevant's cantankerous.
Sturtevant crossed flies with long wings (Grand) and vermillion optics (p) with flies with rudimentary wings (m) and blood-red eyes (P). These traits are X-linked.
For example, to find the distance between P (vermilion optics) and M (long wings), Sturtevant performed crosses between flies that had long wings and vermilion optics and flies that had small wings and scarlet optics. These crosses resulted in Fane flies that either had long wings and cherry-red eyes or long wings and vermilion eyes. Sturtevant then crossed these 2 types of F1 flies and analyzed the offspring for evidence of recombination. Unexpected phenotypes observed in the male F2 progeny from this cantankerous were then examined. (Because very picayune recombination occurs in the male person germ line of Drosophila, merely the female Fane chromosomes are considered for predicting phenotypes [Figure 4].) Sturtevant noted four classes of male flies in this F2 generation, every bit shown in Table 1.
The two additional classes of flies that appeared in this generation (long wings with red eyes and rudimentary wings with vermilion optics) could only be explained by recombination occurring in the female germ line.
| Phenotype | Number of Flies | Nature of Related Gametes |
| Long wings, red eyes | 105 | Recombinant |
| Rudimentary wings, cherry-red eyes | 33 | Nonrecombinant |
| Long wings, vermilion eyes | 316 | Nonrecombinant |
| Rudimentary wings, vermilion eyes | 4 | Recombinant |
| Tabular array 1: Class of male files in the Ftwo generation | ||
Sturtevant and then worked out the order and the linear distances between these linked genes, thus forming a linkage map. In doing and then, he computed the distance in an arbitrary unit he called the "map unit," which represented a recombination frequency of 0.01, or 1%. Afterward, the map unit of measurement was renamed the centimorgan (cM), in award of Thomas Chase Morgan, and information technology is even so used today as the unit of measurement of distances along chromosomes.
In addition to describing the lodge of the genes on the X chromosome of fruit flies, Sturtevant's 1913 paper elucidated a number of other interesting points, including the post-obit:
- The relationship betwixt crossing over and genetic map distance
- The effects of multiple crossover events
- The fact that a first crossover can inhibit a 2nd crossover (a phenomenon called interference, which is described later in this commodity)
Mapping Genes Using Recombination Frequency
To improve understand how Sturtevant arrived as his results, let'southward have a closer look at the procedure he followed. In Figure v, the gray-eosin and xanthous-red flies are the parental lines, and all the alleles for these traits are linked on the X chromosome. Therefore, any gray-red or yellow-eosin male offspring are recombinants. As you tin see, ii recombinants result from the cantankerous. We count simply the male progeny because the males have i X chromosome and say-so volition not obscure whatsoever phenotypes (Robbins, 2000). Of course, crossing over can occur only in the female fruit flies, which have ii X chromosomes. Thus, in this cross, the female Fane gametes provide the parental and recombinant gametes that we observe in the Fii progeny.
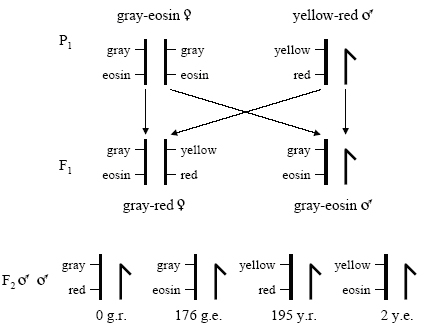
Figure 5: An analogy of Sturtevant'due south cantankerous, showing the chromosomes, illustrates his logic
Sturtevant illustrated the crosses and offspring resulting from a parental strain of grayness-eosin female person flies and xanthous-scarlet male flies. He followed this cross to the F2 generation.
© 2008 Robert J. Robbins. All rights reserved. ![]()
In order to summate the recombination frequency we utilise the following formula:
Substituting the values from our data set up, we arrive at the following:
Therefore, the ii genes are 0.5 map units apart.
Deviations from Expected Results Revealed Genetic Interference
Sturtevant also described the fact that, for genes that were distant from one another, there was a discrepancy in the predicted number of crossovers. For case, the distance between B and M on his map was 57.half dozen. His recombination information using those two genes, withal, did not suggest this distance. Instead, Sturtevant found 260 recombinants in 693 male progeny, which, when plugged into the equation, produced a result of 37.six. How, then, did Sturtevant explain the deviation?
In brusk, Sturtevant realized that double recombination events could occur if genes were far apart. Moreover, non only did Sturtevant's data advise that double-crossing over occurred, but it also suggested that an initial crossover event could inhibit subsequent events by way of a miracle Sturtevant referred to every bit interference.
To sympathise how Sturtevant arrived at this conclusion, accept a look at the information shown in Figure 6 (Sturtevant, 1913). Every bit you can see, Sturtevant examined recombination events between B (body color), CO (two center color genes that were closely linked), and R (rudimentary wings), and compared the frequencies of crossover events. When B and CO did non dissever, Sturtevant noticed that the "gametic ratio," or presence, of CO/R recombinants was approximately ane:two (3,454:vi,972). However, when a crossover betwixt B/CO (N = 60) occurred, there was a much lower likelihood (approximately ane:half dozen.5) of a crossover between CO/R (North = nine). This finding is indicative of interference.
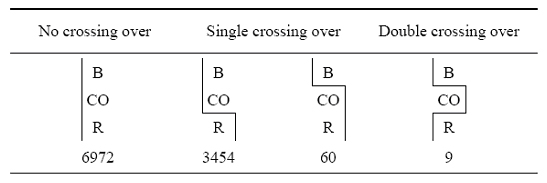
Effigy 6: Information collected by Sturtevant
Number of possible combinations in forms having from 2 to 36 chromosomes in the pre-synaptic cells.
© 2008 Robert J. Robbins. All rights reserved. ![]()
Interference phenomena are even so being studied today, and research has shown that interference can human activity over extremely large distances of the genome. For example, Kenneth J. Hillers and Anne K. Villeneuve recently demonstrated that in Caenorhabditis elegans , interference can actually occur over half the genome of the organism. They demonstrated this past fusing multiple chromosomes together and observing that crossovers still occurred a single fourth dimension (Hillers & Villeneuve, 2003).
Consummate and Incomplete Linkage
When Sturtevant drew his chromosomal map, he placed the C and O genes at the same location because they were ever inherited together (Effigy 3; Sturtevant, 1913). Genes that are so close together on a chromosome that they are always inherited as a single unit show a human relationship referred to as consummate linkage. In fact, ii genes that are completely linked can only be differentiated equally separate genes when a mutation occurs in one of them. There is no other mode to identify genes with consummate linkage from unmarried genes that evidence multiple phenotypes.
On the other hand, the phenomenon known as incomplete linkage occurs when ii genes show linkage with a recombination level greater than 0% and less than 50%. In incomplete linkage, all expected types of gametes are formed, but the recombinant gametes occur less often than the parental gametes.
In improver, if 2 genes are on the same chromosome and are far plenty apart that they undergo recombination at least 50% of the time, the genes are independently assorting and practice non show linkage. Genes independently assort at a distance of fifty cM or more apart. This means that no statistical test would allow researchers to measure linkage.
Finally, linked genes that do not independently assort prove statistical linkage. Statistical linkage is detected as departure from independent assortment that favors the parental gametes. Syntenic genes are genes that are physically located on the aforementioned chromosome, whether or not the genes themselves exhibit linkage (Passarge et al., 1999). Therefore, all linked genes are syntenic, but non all syntenic genes testify genetic linkage.
References and Recommended Reading
Blixt, S. Why didn't Gregor Mendel detect linkage? Nature 256, 206 (1975) doi:10.1038/256206a0 (link to article)
Bridges, C. B. Salivary chromosome maps with a fundamental to the banding of the chromosomes of Drosophila melanogaster. Journal of Heredity 26, 60–64 (1935)
———. A revised map of the salivary gland X chromosome. Journal of Heredity 29, 1113 (1938)
Hillers, K., & Villeneuve, A. Chromosome-broad command of meiotic crossing over in C. elegans. Current Biology 13, 1641–1647 (2003) doi:10.1016/j.cub.2003.08.026
Morgan, T. H. Random segregation versus coupling in Mendelian inheritance. Science 34, 384 (1911)
Passarge, E., et al. Incorrect utilize of the term "synteny." Nature Genetics 23, 387 (1999) (link to article)
Pierce, B. Genetics: A Conceptual Approach. (New York, W. H. Freeman & Co., 2005)
Punnett, R. C. Linkage in the sweet pea (Lathyrus odoratus). Periodical of Genetics thirteen, 101–123 (1923)
———. Linkage groups and chromosome number in Lathyrus. Proceedings of the Purple Gild of London: Series B, Containing Papers of a Biological Character 102 236–238. (1927)
Robbins, R. J. Introduction to sex-express inheritance in Drosophila. Electronic Scholarly Publishing Foundations of Classical Genetics Project. http://world wide web.esp.org/foundations/genetics/classical/thm-10a.pdf (2000) (accessed May 19, 2008)
Sturtevant, A. H. The linear arrangement of six sex-linked factors in Drosophila, as shown past their mode of association. Journal of Experimental Zoology 14, 43–59 (1913)
Weiner, J. Time, Honey, Memory: A Great Biologist and His Quest for the Origins of Behavior (New York, Random House, 1999)

Source: http://www.nature.com/scitable/topicpage/thomas-hunt-morgan-genetic-recombination-and-gene-496
Posted by: alcarazliplet.blogspot.com
0 Response to "How To Draw A Genetic Map"
Post a Comment